Cooperative benefit for the combination of rapamycin and imatinib in tuberous sclerosis complex neoplasia
Vascular Cell. 2012;
Received: 18 April 2012 | Accepted: 18 June 2012 | Published: 5 July 2012
Vascular Cell ISSN: 2045-824X
Abstract
Tuberous sclerosis (TS) is a common autosomal-dominant disorder characterized by tumors of the skin, lung, brain, and kidneys. Monotherapy with rapamycin however resulted in partial regression of tumors, implying the involvement of additional pathways. We have previously implicated platelet-derived growth factor-BB in TS-related tumorigenesis, thus providing a rationale for a combination of mTOR/PDGF blockade using rapamycin and imatinib. Here, we test this combination using a well-established preclinical model of cutaneous tumorigenesis in TS, tsc2ang1 cells derived from a skin tumor from a mouse heterozygous for tsc2. Treatment of tsc2ang1 cells with a combination of rapamycin and imatinib led to an inhibition of proliferation compared with either vehicle treatment or treatment with rapamycin or imatinib monotherapy. Combination therapy also led to a decrease in Akt activation. Potent
Introduction
TS is a common autosomal-dominant disorder characterized by the development of tumors of the brain, kidney, skin and lung. The disorder is characterized by mutations or deletions in one of two large genes, hamartin (
The signaling pathways implicated in TS are complex. A hotspot for mutations in tsc2 involves regions implicated in controlling rheb, although multiple other signaling pathways have also been linked to TS-related neoplasia, including mTORC1, notch, p42/44 MAP kinase, NFkB, and Akt [2–11]. Tuberin (
We have previously demonstrated that platelet-derived growth factor β receptor (PDGFRβ) is present and active in human and murine TS lesions [15–17]. Other groups have demonstrated an inverse relationship between mTOR activation and PDGFRβ levels in TS-derived cells [18]. Therefore, we reasoned that mTOR blockade might be compensated for by PDGF activation in vitro and
Results
Rapamycin and imatinib treatment inhibits TSC2 ang1 cell proliferation
To assess if rapamycin and imatinib combination was superior to either agent alone, we assessd their effects on the proliferation of Tsc2 ang1 cells. Treatment of cells for 72 h with rapamycin (5nM) or imatinib (10 μM) alone reduced the level of proliferation significantly compared with control (p = 0.0006 for imatinib vs control, p = 0.0076 for rapamycin vs control). However, combined treatment with with both agents led to significantly better inhibition of proliferation (Figure 1a), compared with control (p = 0.0002). Combination of drugs was significantly better than rapamycin alone (p = 0.0243), while imatinib vs combination did not reach significance at p less than 0.05. Experiments were done in triplicates for reproducibility.
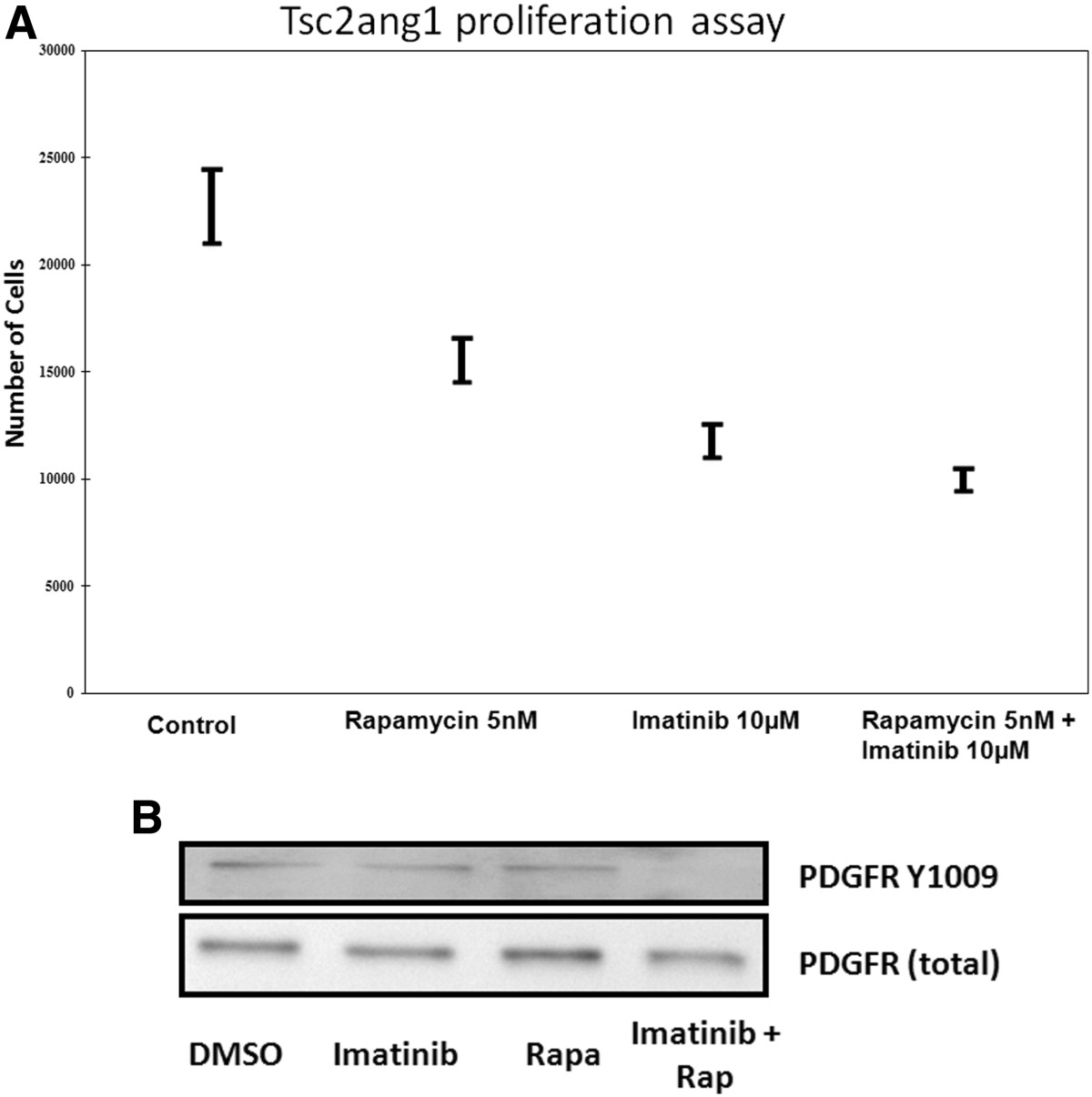
Figure 1
Figure 1 caption
(A) Effect of rapamycin, imatinib or combination of imatinib and rapamycin on tsc2ang1 cell line proliferation. Cells were treated with vehicle control (DMSO), rapamycin (5nM), imatinib (10 μM) or imatinib (10 μM) + rapamycin (5nM) (shown along X-axis) for 24 h and counted using a Coulter counter. Treatment at each concentration was performed in triplicate. The Y-axis represents cell number. (B) Downregulation of Phospho-PDGFR-β by combinational treatment; Tsc2ang1 cells were treated with DMSO, rapamycin5nM, imatinib10uM or rapamycin5nM + imatinib10uM for 24 hours prior to harvest. Combination rapamycin + imatinib treatment inhibits the levels of phospho- PDGFR-β (Tyr 1021).
Combination of rapamycin and imatinib inhibits PDGFRβ activation inTsc2 ang1 cells
In view of the effects of rapamycin and imatinib combination on Tsc2Ang1 cell proliferation together with previous findings that mTOR inhibitor treatment of TS is associated with increased PDGFRβ levels, we examined the effects of imatinib and rapamycin or a combination of these on PDGFRβ expression. Experiments were done in triplicates for reproducibility. We tested imatinib (10 μM), rapamycin (5nM) or their combination on the levels of phosphorylated PDGFRβ (at Tyr-1009) in Tsc2ang1cells. Combined treatment led to downregulation of PDGFRβ phosphorylation, (Figure 1b).
Treatment with combination of rapamycin and imatinib down regulates phosphoAkt473 protein levels in Tsc2ang1 cells
To verify the effect of imatinib and rapamycin, as well as combination therapy on the activation of Akt, we treated Tsc2ang1 cells with imatinib (10 μM), rapamycin (5 nM) or their combination and the levels of Akt and p Akt were measured by densitometry scanning of the signals on western blot membrane. Total levels of Akt were unchanged by treatment and served as loading controls. We found first that overall, treatment with drug led to significantly decreased phospho Akt compared to vehicle controls (p = 0.0002, general linear modeling). In addition, we found that treatment with rapamycin and imatinib reduced the levels of pAkt (Figure 2) and a significant reduction in the levels of pAkt (p <0.0002) in imatinib + rapamycin treated cells compared to vehicle control. The mean of combination treatment versus imatinib alone or rapamycin alone also decreased. However, significance was only observed when the combination treatment was compared to rapamycin alone (p < 0.0243) and not when compared to imatinib alone (p < 0.4273).
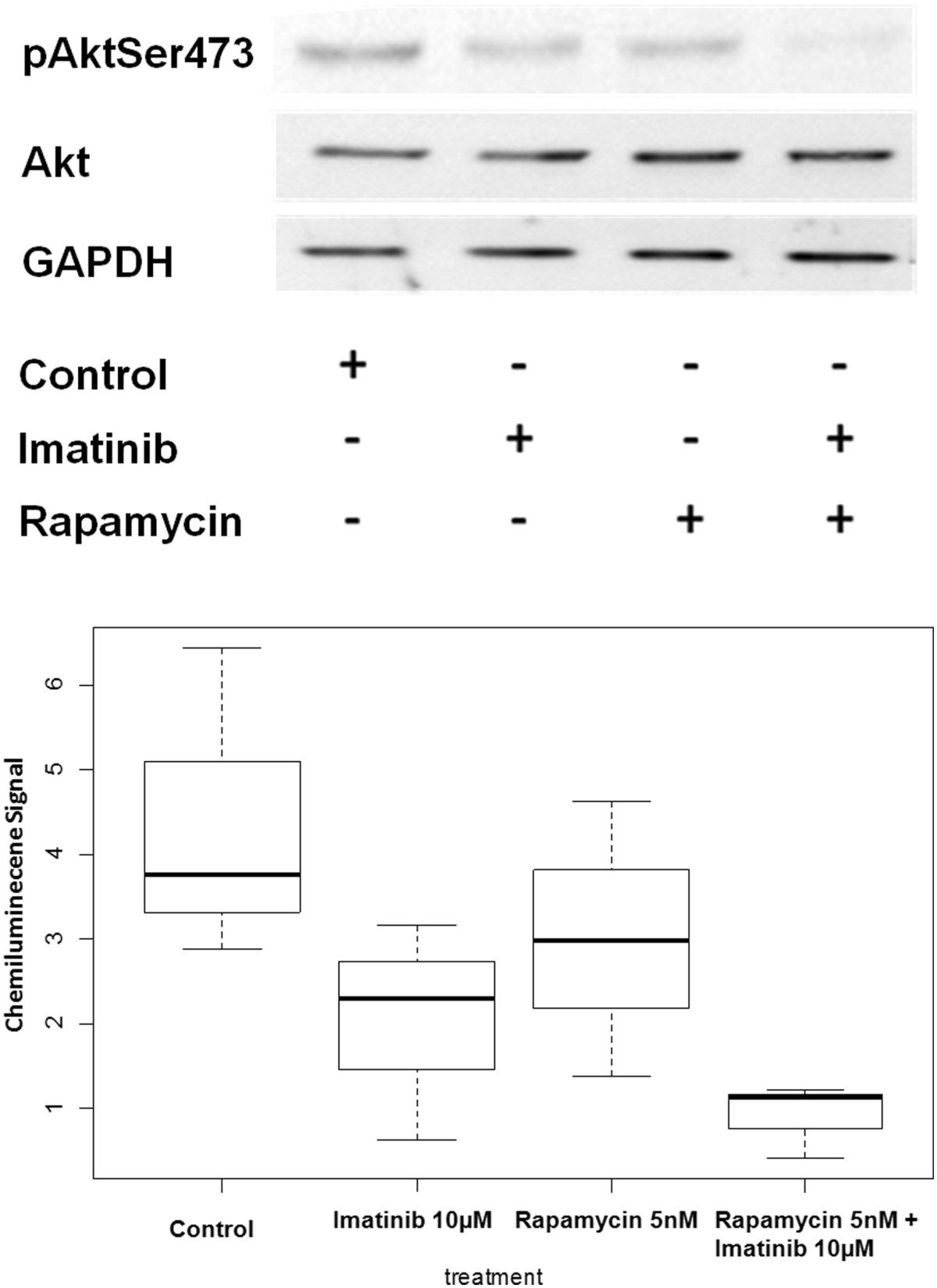
Figure 2
Figure 2 caption
blot analysis using Akt and pAkt were conducted on Tsc2 ang1 cells treated with treated with DMSO, rapamycin5nM, imatinib10uM or rapamycin5nM + imatinib10uM for 24 hours. Cells were lysed and analyzed by using antibodies specific for the total Akt and pAkt473. Rapamycin + imatinib treatment downregulates pAkt levels (p <0.0002). GAPDH was used as the loading control by using monoclonal anti-GAPDH antibody. All treatment groups were significant when compared to the vehicle control, Imatinib 10nM (p < 0.0006); Rapamycin 5nM (p < 0.0076); Rapamycin 5nM + Imatinib 10nM (p < 0.0002). However, the combinational therapy only show significance against Rapamycin at 5nM (p < 0.0243) and not Imatinib at 10nM (p < 0.4273).
Matrix metalloproteinase analysis
Cell migration and invasion are fundamental components of tumor cell invasion and neovascularization. Matrix metalloproteinases (MMPs) are essential for successful cell migration through extracellular matrix. In order to determine the levels of MMPs in our samples, conditioned media from equivalent numbers of imatinib- or rapamycin-treated cells along with that of control cells were collected and quantitatively analyzed using monospecific ELISAs. MMP-2 levels in conditioned media from tsc2ang1 cells treated with either imatinib (p = .0243), rapamycin or combination treatment with rapamycin and imatinib (p = .0092) were significantly decreased compared to that of controls (Figure 3). However, MMP-2 levels in the conditioned media of cells treated with rapamycin (p = .0744) were not significantly decreased compared to that of controls consistent with previous reports [20]. Differences between rapamycin alone and combination therapy trended toward significance (p = 0.0744), and between imatinib alone and combination (p = 0.329), indicating that imatinib has a more potent effect on MMP-2 activity than rapamycin. Moreover, no significant difference was observed in MMP-9 levels between the different treatment groups and controls (data not shown).
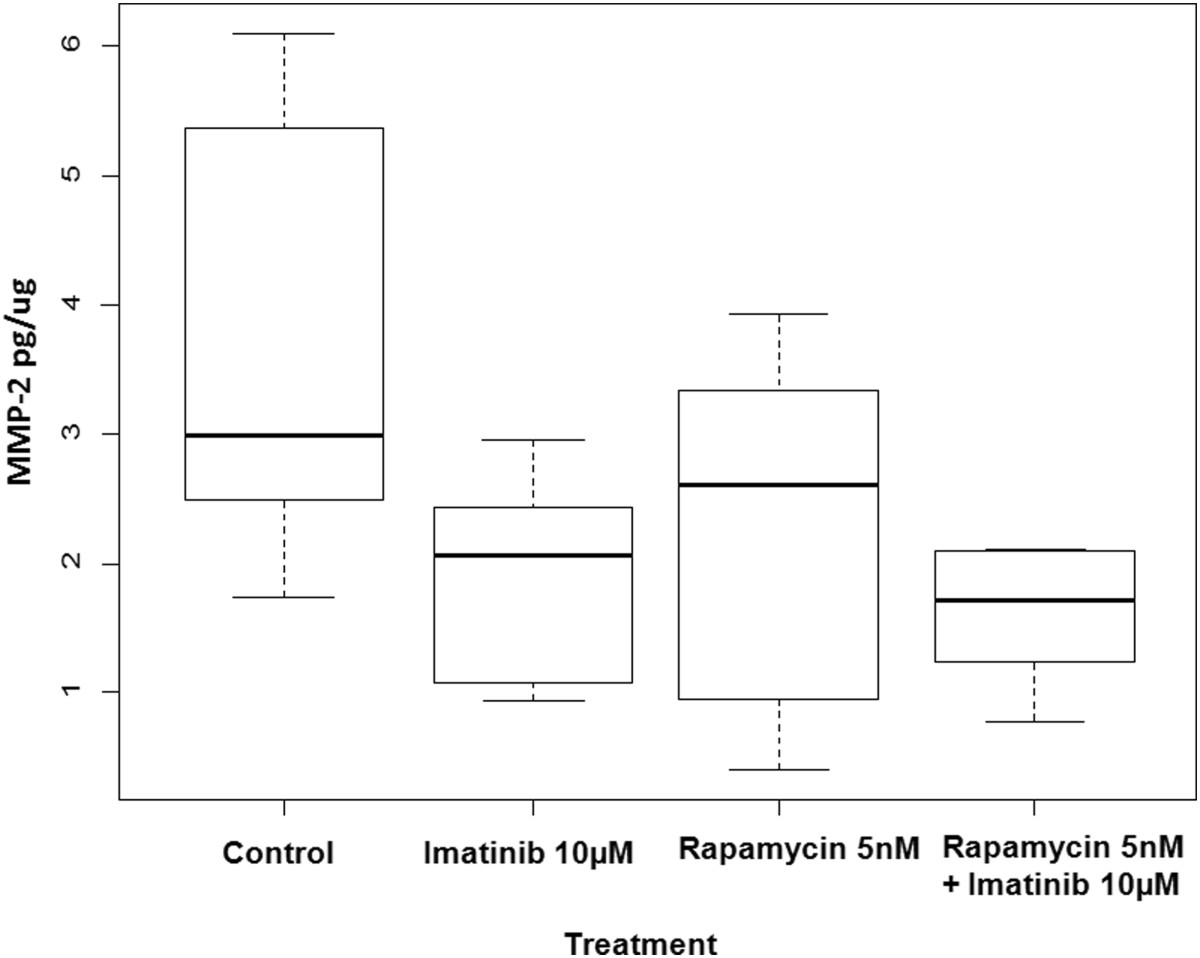
Figure 3
Figure 3 caption
MMP-2 ELISA analysis of conditioned media from tsc2ang1 cells treated with either rapamycin, imatinib or the combination for 24 hours resulted in a decrease in the amount of MMP-2 produced in comparison to controls. MMP-2 levels in the tsc2ang1 cells treated with imatinib only and imatinib + rapamycin were significantly decreased compared to that of controls.
In vivo tumorigenesis
Given the effects of combined treatment with imatinib and rapamycin on cell proliferation as well as the ability of this combination to reduce the levels of phospho-PDGFRβ and phosphor-Akt, we assessed the effects of this combination on tsc2ang1 tumorigenesis
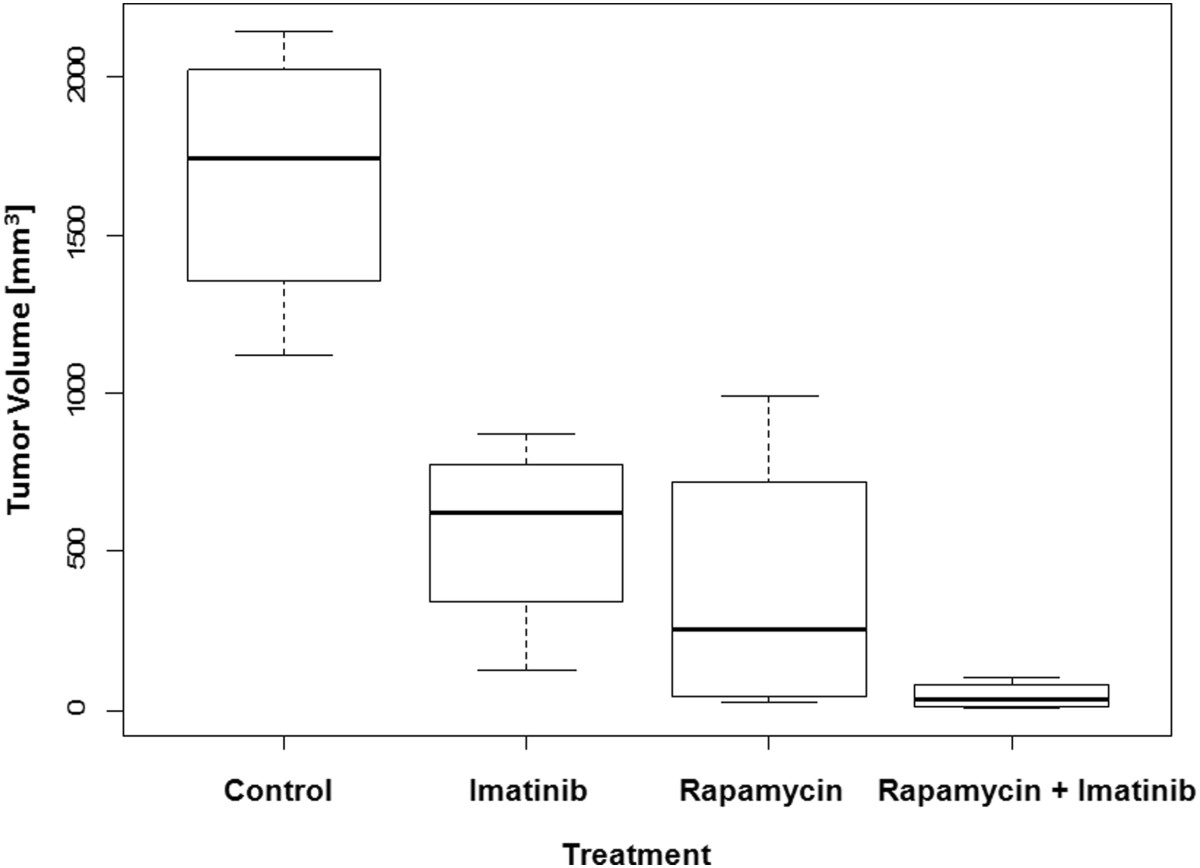
Figure 4
Figure 4 caption
Effect of rapamycin and imatinib ongrowth of tsc2ang1 xenografts in nude mice. The y axis represents tumor volume.
Discussion
TS is a multisystem disorder characterized by benign or malignant neoplasia, as well as autism and seizures. No highly effective and long lasting therapy exists for TS neoplasia. Renal lesions such as angiomyolipomas may cause massive bleeding and compromise renal function, often requiring a kidney transplant. Lymphangiomyomatosis, a neoplastic complication most commonly seen in young women, can only be cured by lung transplantation. Deforming skin lesions cause significant psychological distress [21]. Finally, brain tumors such as subependymal giant cell astrocytomas, can cause morbidity and mortality, and multiple tubers results in refractory seizures and exacerbation of mental retardation. Thus, urgent therapy is required for TS. Here, we demonstrate in a validated preclinical model a nearly complete tumor inhibition with a combination of two FDA-approved drugs (rapamycin and imatinib) targeting two distinct signaling pathways (mTORC1 and PDGFRβ, respectively) implicated in TS-associated neoplasia.
The lack of total blockade by rapamycin is consistent with the incomplete clinical response of tumors to systemic rapamycin, implying that either additional pathways are already present in these tumors, or are induced by rapamycin monotherapy. High among the candidates are the PDGFRβ mediated signal pathways, especially since mTORC1 activation and PDGFRβ signaling have been shown to have an inverse relationship [18].
Treatment with combination of rapamycin and imatinib led to downregulation of PDGFRβ in tsc2ang1 cells, in a c-cbl-independent manner (data not shown). In vitro proliferation assays demonstrated an additive effect of rapamycin and imatinib on tsc2ang1 cells.
Rapamycin has been used as monotherapy in patients with TS, resulting in benefit only as long as the patients are exposed to the drug [12]. Our findings that rapamycin therapy alone does not address PDGFβ signaling, as well as modest efficacy against Akt activation are potential reasons for the failure of rapamycin as monotherapy in solid tumors. In highly malignant tumors, mTORC1 inhibition has shown to cause a paradoxical activation of Akt, in part through activation of mTORC2, which is an Akt kinase [22, 23]. We demonstrate that the combination of rapamycin and imatinib blocks Akt activation compared with monotherapy, and likely accounts for the decrease in tumor volume. Our findings demonstrate the following. Combination therapy of TS relevant cells with rapamycin and imatinib results in downregulation of PDGFRβ and Akt signaling. This decrease is independent of c-cbl-mediated degradation. Finally, the combination of rapamycin and imatinib has at least additive activity against the tsc2ang1 model of TS
Materials and methods
Generation of murine model of TS
Tsc2ang1 (ATCC CRL 2620) is a murine cell line derived from a cutaneous sarcoma that arose in the extremity of a mouse heterozygous for tsc2; these mice develop cutaneous sarcomas at a frequency of 10 to 15%. The cells were cultured in complete DMEM medium supplemented with 10% FBS (Sigma Aldrich, St Louis, MO).
In vitro proliferation assay
10,000 tsc2ang1 cells per well were plated in 24-well dishes in triplicate. The next day, fresh medium containing the compounds or vehicle controls was added. Cells were incubated at 37° C for 24 h, and cell number was determined using a Coulter Counter (Hialeah, FL).
Western blot analysis
Lysates of Tsc2ang1 cells treated with vehicle or indicated drugs were prepared in NP-40 lysis buffer (1% NP-40, 150 mmol/L NaCl, 10% glycerol, 20 mmol/L HEPES, 1 mmol/L phenylmethylsulfonyl fluoride, 2.5 mmol/L EDTA, 100 μmol/L Na3VO4, and 1% aprotinin). Protein concentration in cleared lysates was determined using an Eppendorf BioPhotometer. Samples were resolved using SDS-PAGE (National Diagnostics) and transferred to nitrocellulose membranes. The membranes were blocked with 5% nonfat dry milk in 10 mmol/L Tris/0.1% Tween 20/100 mmol/L NaCl and incubated with primary antibodies followed by horseradish peroxidase–conjugated secondary antibody. The immunoreactive bands were visualized by enhanced chemiluminescence (Amersham Biosciences). The antibodies used were: Phospho-PDGFR- β antibody(Tyr 1021) (Cell signaling Laboratories); Akt antibody (9272) (Cell signaling Laboratories), pAKT (4058) antibody (Cell signaling Laboratories) monoclonal anti-GAPDH antibody (Santa Cruz L-18 S-48167) was used as a loading control.
Matrix metalloproteinase analysis
The presence of MMPs in conditioned media samples was determined using MMP ELISA Quantikine Kits (R&D Systems, Inc.). Specimens, standards and reagents were prepared according to manufacturer's instructions. Protein concentration was determined via the Bradford method using bovine serum albumin as the standard as described previously [24].
In vivo tumor growth
To test the
Statistics
One way ANOVA, and non parametric test were preformed for the tumor volume statistics. We did parametric analysis when the conditions for ANOVA were satisfied and in case where conditions are not satisfied and if the variables are not normally distributed, we conducted corresponding non parametric test and opted to present results for Wilcoxon test.
Ethical approval
Animals studies were performed in compliance with the Emory IACUC.
Acknowledgements
JLA was supported by the grant RO1 AR47901and P30 AR42687 Emory Skin Disease Research Core Center Grant from the National Institutes of Health, a Veterans Administration Hospital Merit Award, as well as funds from the Rabinowitch-Davis Foundation for Melanoma Research and the Betty Minsk Foundation for Melanoma Research. JLA was also funded by Robert Margolis Liposarcoma Research Fund. HB was supported by NIH grants CA87986, CA105489, CA 116552 and CA99163 and MAM and ASC were supported by NIH grant P01 CA045548.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Authors’ original file for figure 1
Authors’ original file for figure 2
Authors’ original file for figure 3
Authors’ original file for figure 4
Authors’ original file for figure 5
Authors’ original file for figure 6
References
- Inflammatory processes in cortical tubers and subependymal giant cell tumors of tuberous sclerosis complex. Epilepsy Res. 2008;78:7-21.
- Mutation in TSC2 and activation of mammalian target of rapamycin signalling pathway in renal angiomyolipoma. Lancet. 2003;361:1348-1349.
- Platelet-derived growth factor-induced p42/44 mitogen-activated protein kinase activation and cellular growth is mediated by reactive oxygen species in the absence of TSC2/tuberin. Cancer Res. 2005;65:10881-10890.
- Tuberin regulates p70 S6 kinase activation and ribosomal protein S6 phosphorylation. A role for the TSC2 tumor suppressor gene in pulmonary lymphangioleiomyomatosis (LAM). J Biol Chem. 2002;277:30958-30967.
- The evolutionarily conserved TSC/Rheb pathway activates Notch in tuberous sclerosis complex and Drosophila external sensory organ development. J Clin Invest. 2010;120:93-102.
- Rhebbing up mTOR: new insights on TSC1 and TSC2, and the pathogenesis of tuberous sclerosis. Cancer Biol Ther. 2003;2:471-476.
- Measurements of TSC2 GAP Activity Toward Rheb. Methods Enzymol. 2005;407:46-54.
- Regulation of TSC2 by 14-3-3 binding. J Biol Chem. 2002;277:44593-44596.
- Phosphorylation and functional inactivation of TSC2 by Erk implications for tuberous sclerosis and cancer pathogenesis. Cell. 2005;121:179-193.
- Akt regulates growth by directly phosphorylating Tsc2. Nat Cell Biol. 2002;4:658-665.
- Rheb is a direct target of the tuberous sclerosis tumour suppressor proteins. Nat Cell Biol. 2003;5:578-581.
- Sirolimus for angiomyolipoma in tuberous sclerosis complex or lymphangioleiomyomatosis. N Engl J Med. 2008;358:140-151.
- Rapamycin causes regression of astrocytomas in tuberous sclerosis complex. Ann Neurol. 2006;59:490-498.
- TSC2 regulates VEGF through mTOR-dependent and -independent pathways. Cancer Cell. 2003;4:147-158.
- Functional tyrosine kinase inhibitor profiling: a generally applicable method points to a novel role of platelet-derived growth factor receptor-beta in tuberous sclerosis. Am J Pathol. 2002;161:781-786.
- Transgenic expression of dominant negative tuberin through a strong constitutive promoter results in a tissue-specific tuberous sclerosis phenotype in the skin and brain. J Biol Chem. 2005;280:5870-5874.
- Malignant transformation of human cells by constitutive expression of platelet-derived growth factor-BB. J Biol Chem. 2005;280:13936-13943.
- PDGFRs are critical for PI3K/Akt activation and negatively regulated by mTOR. J Clin Invest. 2007.
- Down-regulation of MMP-2 expression due to inhibition of receptor tyrosine kinases by imatinib and carboplatin in HNSCC. Oncol Rep. 2011;25:1145-1151.
- Rapamycin-insensitive up-regulation of MMP2 and other genes in tuberous sclerosis complex 2-deficient lymphangioleiomyomatosis-like cells. Am J Respir Cell Mol Biol. 2010;42:227-234.
- MCP-1 overexpressed in tuberous sclerosis lesions acts as a paracrine factor for tumor development. J Exp Med. 2005;202:617-624.
- mSin1 is necessary for Akt/PKB phosphorylation, and its isoforms define three distinct mTORC2s. Curr Biol. 2006;16:1865-1870.
- Activation of Akt and eIF4E survival pathways by rapamycin-mediated mammalian target of rapamycin inhibition. Cancer Res. 2005;65:7052-7058.
- Urinary biomarkers predict brain tumor presence and response to therapy. Clin Cancer Res. 2008;14:2378-2386.
- Tris (dibenzylideneacetone) dipalladium, a N-myristoyltransferase-1 inhibitor, is effective against melanoma growth in vitro and in vivo. Clin Cancer Res. 2008;14:5743-5748.

The present article has been published in Vascular Cell journal by Publiverse Online S.R.L.