Ageing and microvasculature
Vascular Cell. 2014;
Received: 13 March 2014 | Accepted: 15 August 2014 | Published: 16 September 2014
Vascular Cell ISSN: 2045-824X
Abstract
A decline in the function of the microvasculature occurs with ageing. An impairment of endothelial properties represents a main aspect of age-related microvascular alterations. Endothelial dysfunction manifests itself through a reduced angiogenic capacity, an aberrant expression of adhesion molecules and an impaired vasodilatory function. Increased expression of adhesion molecules amplifies the interaction with circulating factors and inflammatory cells. The latter occurs in both conduit arteries and resistance arterioles. Age-related impaired function also associates with phenotypic alterations of microvascular cells, such as endothelial cells, smooth muscle cells and pericytes. Age-related morphological changes are in most of cases organ-specific and include microvascular wall thickening and collagen deposition that affect the basement membrane, with the consequent perivascular fibrosis. Data from experimental models indicate that decreased nitric oxide (NO) bioavailability, caused by impaired eNOS activity and NO inactivation, is one of the causes responsible for age-related microvascular endothelial dysfunction. Consequently, vasodilatory responses decline with age in coronary, skeletal, cerebral and vascular beds. Several therapeutic attempts have been suggested to improve microvascular function in age-related end-organ failure, and include the classic anti-atherosclerotic and anti-ischemic treatments, and also new innovative strategies. Change of life style, antioxidant regimens and anti-inflammatory treatments gave the most promising results. Research efforts should persist to fully elucidate the biomolecular basis of age-related microvascular dysfunction in order to better support new therapeutic strategies aimed to improve quality of life and to reduce morbidity and mortality among the elderly patients.
Keywords
Endothelial cells Smooth muscle cells Endothelial dysfunction Nitric oxide Vascular remodelling Organ-specific ageingIntroduction
Vascular ageing is associated with both structural and functional changes that can take place at the level of the endothelium, vascular smooth muscle cells and the extracellular matrix of blood vessels [1]. Age, hypertension, diabetes, smoking and plasma low density lipoprotein cholesterol level are determinant risks of arterial stiffness [2, 3]. A relevant age-related vascular change is a progressive myointimal thickening [4, 5]. Similarly to that observed in large vessels, age-related increase of microvascular tone leads to a progressive myogenic hypertrophic remodelling of small arteries, due to the increased distending pressure acting perpendicularly on the vascular wall [6]. Microvascular alterations play an important role in ageing-associated end-organ damage [7]. In fact, microcirculation provides the interface for tissue delivery of oxygen and nutrients, removal of waste products and carbon dioxide, transvascular exchange and fluid economy [8]. Therefore, cell survival depends on adequate microvascular perfusion [8]. The architecture and the biophysical behavior of flowing blood strongly influence microvascular function. Morphologically, the microcirculation is constituted from vessels <300 μm in diameter [8]. Therefore, it includes arterioles, capillaries, and venules (Figure 1). Alternatively, a physiological definition based on vessel function rather than diameter or structure has been proposed [9]. By this definition, vessels that respond to an increase of pressure by a myogenic reduction in lumen diameter are considered part of the microcirculation [9]. Consequently, besides endothelial cells, also vascular smooth muscle cells (VSMCs) and pericytes must be included in the microvascular cell population. Although the primary function is to optimise the nutrient and oxygen supply, microcirculation is relevant in order to avoid large hydrostatic pressure fluctuations causing disturbances in capillary exchange and an overall peripheral vascular resistance [10]. An important role in regulating tissue fluid balance and in maintaining osmotic and hydrostatic pressures is played by the lymphatic system (Figure 1), that comprises a one-way transport for fluid and proteins by collecting them from the interstitial space and returning them to the blood circulation [11]. This review focuses the attention on the biomolecular and pathophysiological mechanisms underlying age-related microvascular alterations and the importance of new therapies to prevent end-organ damage associated with microvascular dysfunction.
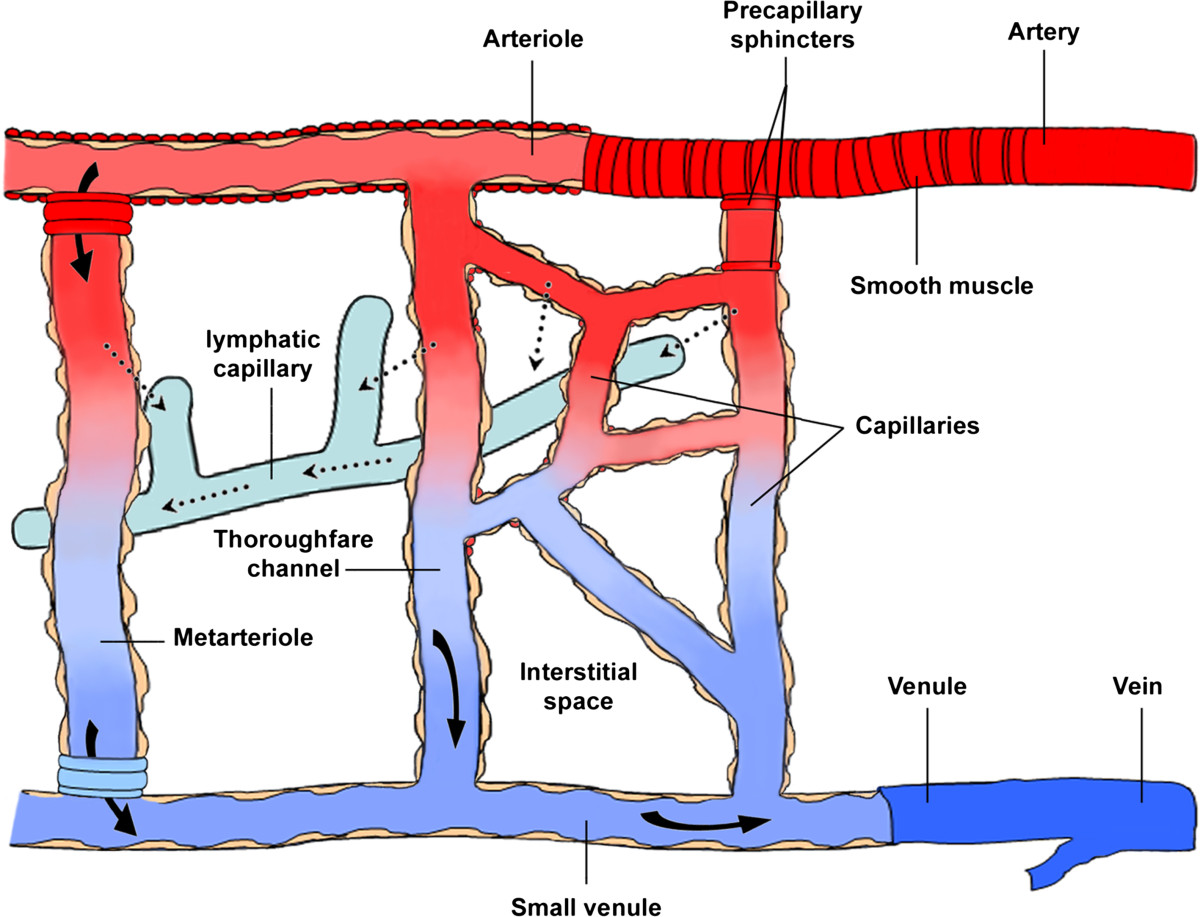
Figure 1
Figure 1 caption
Schematic representation of microcirculation components. The microcirculation is a network of small blood vessels, including arterioles, venules and capillaries. Blood flows from the arteries into the arterioles and then pass into the venules across true capillaries or throughfare channels and metarterioles (arteriovenous bypass). The precapillary sphincter, made of smooth muscle cells, controls blood flow into the true capillaries. As blood travels through the capillaries, plasma proteins and fluid enter the interstitial space according to hydrostatic and osmotic pressure gradients. Most of the fluid is reabsorbed into the post-capillary venules, while a fraction enters to the lymphatic circulation for its return to the blood circulation.
Biomolecular mechanisms involved in age-related microvascular dysfunction
Nitric oxide
In mammals, nitric oxide (NO) is produced by a family of enzymes, named nitric oxide synthases (NOSs), that catalyse the production NO from L-arginine. NO is an important cellular signalling molecule that regulates vasodilatation, insulin secretion, airway tone, and peristalsis, and is involved in angiogenesis and neural development [29]. The family of enzymes NOS comprises three isoforms: neuronal NOS (nNOS/NOS1), inducible NOS (iNOS/NOS2) and endothelial NOS (eNOS/NOS3) [29] eNOS constitutively produces NO in endothelial cells and physiologically contributes to the control of vascular tone. Instead iNOS is activated by bacterial lipopolysaccharide, cytokines, and other inflammatory agents, determining an abnormal production of NO. Due to its affinity to protein-bound iron, NO can inhibit key enzymes that contain iron in their catalytic centers. These include iron–sulfur cluster-dependent enzymes (complexes I and II) involved in mitochondrial electron transport, ribonucleotide reductase (the rate-limiting enzyme in DNA replication), and
As discussed above, microvascular dysfunction is mainly induced by the over-production and release of O2-, which cause NO breakdown. In fact, NO inactivation is due to its reaction with O2- to form the potent oxidant peroxynitrite (ONOO−) [30]. This compound can cause oxidative damage, nitration, and S-nitrosylation of biomolecules including proteins, lipids, and DNA single-strand breakage following the poly-ADP-ribose polymerase (PARP) activation [31–33]. The increase of nitration was demonstrated in the sarcoplasmic reticular Ca-ATPase isolated from the skeletal muscle of old rats [34]. The scavenging of NO by O2- was also demonstrated in coronary microvascular endothelial cells of old rats, in which the reduction of eNOS expression was accompanied with an increased O2- production and attenuated vasodilator responses [35]. Coronary arterioles of aged rats displayed an increased iNOS activity and ONOO− production, as well as a decreased eNOS expression [36]. The same alterations have been also described in elderly [36].
Moreover, oxidative stress can convert eNOS from a NO-producing enzyme to an enzyme that generates O2-. This process is named eNOS uncoupling. Mechanisms implicated in eNOS uncoupling include oxidation of the critical NOS cofactor BH4, depletion of L-arginine, and accumulation of endogenous methylarginines [29].
Age-related signal alterations in vascular cells
It has been demonstrated that endothelin-1 and angiotensin II (potent vasoconstrictors) pathways are involved in age-related endothelial oxidative stress [18]. In particular, ageing induced endothelin-1 overexpression, resulting in vascular remodelling and endothelial dysfunction in mice [37]. In addition, it has been reported the involvement of endothelin-1 in eNOS downregulation in pulmonary artery endothelial cells of fetal porcine [38]. As concerning angiotensin II, it has been documented that in ageing its overexpression caused vascular senescence by mitochondrial and NADPH-dependent superoxide generation [18]. This mechanism was attenuated by mitochondrial electron transport chain or angiotensin type 1 receptor inhibitors [39, 40]. Moreover, the infusion in rats of angiotensin II induced microvascular lesions in various vascular beds that resemble arteriolosclerosis [41]. The blocking of nitric oxide synthesis also induced renal microvascular disease [42].
It is well known that angiogenesis and wound healing are reduced with ageing [43]. In fact, vascular endothelial growth factor (VEGF)-induced angiogenesis is attenuated in aged rats and rabbits [44, 45]. In aged mice and in cultured human microvascular endothelial cells aged by progressive passaging, the expression of the tissue inhibitor of metalloproteinase-2 (TIMP-2) is increased [46], and correlated with an attenuated capacity of endothelial cells to degrade extracellular matrix, a process required for angiogenesis [46].
Taken together these findings suggest the existence of a complex biomolecular mechanism involved in age-related vascular dysfunction that leads to oxidative stress, vascular remodelling and endothelial dysfunction. This altered signalling, in endothelial cells, causes the activation of NF-kB and a consequent abnormal gene transcription, including the enhancement of cellular adhesion molecule expression, such as intercellular adhesion molecule-1 (ICAM-1), vascular cell adhesion molecule-1 (VCAM-1), E-selectin, and inflammatory cytokine secretion [47–49]. This process determines the leukocyte recruitment and extravasation as also demonstrated in the vascular wall of aged rabbits [50]. A schematic representation of age-related biomolecular alterations in microcirculation is reported in Figure 2.
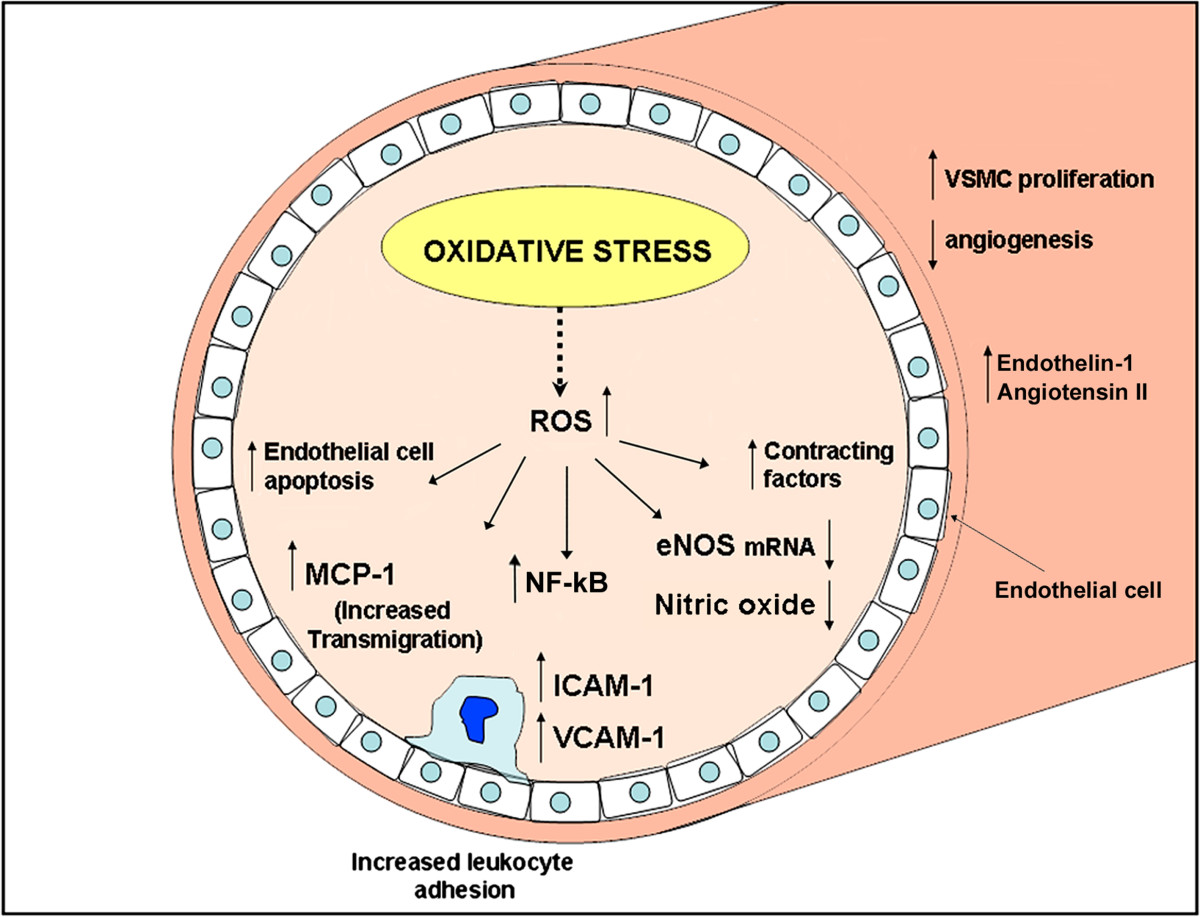
Figure 2
Figure 2 caption
Schematic representation of biomolecular changes in age-related microvascular dysfunction. Oxidative stress plays a pivotal role in endothelial and myocitic impaired function.
Structural and functional microvascular alterations involved in ageing
Endothelial cells
As a consequence of the alteration in the expression and/or activity of eNOS, upregulation of iNOS, and increased formation of ROS and ONOO−, endothelial cells undergo to cumulative DNA damage that promotes senescence and apoptosis [52]. As described above, the age-related decline of endothelial function becomes manifest through a reduced regenerative and angiogenic capacity, and an altered expression of adhesion molecules regulating the interaction of circulating factors with immune system cells [53, 54].
The attenuated capacity of the endothelium to regenerate is partially a consequence of an impaired secretion of and/or sensitivity to growth factors [55]. Recently, the regeneration of the endothelium by bone marrow-derived circulating progenitor cells has gained particular attention, because the number of circulating endothelial progenitor cells (EPCs) decreases with age and is thought to reflect the attenuated mobilization of these cells from the bone marrow [56]. Moreover, EPCs from older subjects have a reduced capacity to engraft [57]. Some studies suggest that the regenerated endothelium is functionally impaired [57] and exhibits an increased uptake of modified low-density lipoprotein (LDL) and decreased NO production [58]. For example, as documented in aged rats, important structural changes of brain capillaries were found: thickening of the basal lamina and the thinning of endothelial cells [59]. Some suggest that this phenomenon is due to a loss of endothelial cells together with a lengthening of the remaining ones to allow nutrients to diffuse [60]. Mophological alteration of aged endothelium was observed also in sinusoids of human aged liver, where thickening of the sinusoidal endothelium was associated with the deposition of basal lamina and collagen [61]. In the kidney of aged rats the number of proliferating endothelial cells was decreased compared with young rats. In addition, VEGF expression strongly decreased with ageing in the endothelium of the outer and inner medulla, suggesting a reduced angiogenic activity [62].
Smooth muscle cells and pericytes
As discussed above, in ageing, upregulation of pro-oxidants and downregulation of antioxidants results in an imbalance leading to ROS increase [63–65] and to the development of vascular dysfunction in both animal models and in humans [66]. In old rats, a significant increase in O2- was observed in the vascular wall [67], and was associated with an increase in NAD(P)H oxidase activity [36, 64, 68–70]. It has been also reported that Angiotensin II pathway plays an important role in age-related smooth muscle cell oxidative stress by eliciting NAD(P)H oxidase activity [71]. In fact, Angiotensin II stimulation induced the NAD(P)H oxidase-dependent O2- production, stimulating NF-κB signalling in senescent VSMCs [72]. Similarly to endothelial cells, VSMCs of old rats in response to cytokines showed higher ICAM-1 level compared with newborn rats [73]. VSMCs can also induce the activity of iNOS through the NF-kB pathway under inflammatory conditions [64], as also reported in aged Macaca mulatta, rats [64, 74] and mice [75]. As already reported, vascular ageing is also associated with a progressively reduced NO bioavailability. Since VSMCs are important targets for endothelium-derived NO, this reduction causes an impairment of endothelium-dependent vasodilation [76]. In addition, the
Pericytes, the mural cells on capillaries, play an important role in vessel stabilisation, by regulating endothelial cell proliferation and preventing capillary withdrawal [83–85]. Alterations in these cells with ageing also might contribute to the development of age-related morphological and physiological abnormalities of the microvasculature. In fact, microvascular ageing is characterised by changes in peripheral capillaries, including vessel broadening, and thickening of the basement membrane, as well as altered length and orientation of desmin filaments in pericytes [86]. These changes can determine a reduced pericyte–endothelial cell contact, destabilisating capillaries [86]. In addition, a reduction in pericyte number in aged capillaries was also reported [87]. In the brain capillaries of elderly the decrease in pericyte coverage was reported [88]. It has been also documented that in the retina of old rats, ageing induced the broadening of peripheral capillaries and terminal venules, as well as thickening of basement membranes [86]. In the retina of old rats was reported a shift from a pericyte phenotype toward an arteriolar smooth muscle cell–like phenotype. It was associated with an increase in calponin labelling of arterioles, thickness of basement membranes, and increased focal adhesions in arteriolar walls [86]. Moreover, in skeletal muscle of old mice, the muscular regenerative capacity of pericytes is limited, and they produce collagen and contribute to fibrous tissue depositing [89].
Lymphatic vessel alterations
Lymphatic system begins when the plasma fluid and proteins, that are forced out by arterial capillaries into the interstitial space (Figure 1), are collected into the lymphatic capillaries, which are freely permeable to macromolecules [90]. So, the main function of lymphatic system is to maintain osmotic and hydrostatic pressures within the tissue space. It consists of capillaries (10-60 μm in diameter) that drain lymph into the collecting vessels that contain also smooth muscle. The fluid pass through several clusters of lymph nodes and then into larger trunks, which in turn lead into the ducts, that return lymph back into the bloodstream [11].
Spontaneous contractions of smooth muscle cells in the wall of lymphatic vessels are necessary to maintain effective lymph flow whereas proper functioning of lymphatic endothelial cells is necessary to regulate lymphatic contractility [91]. The basic self-regulatory mechanisms controlling lymph flow in lymphatic vessels is realised through the sensitivity of their muscle cells to levels of stretch and of their endothelial cells to levels of the shear stress [91]. Nitric oxide plays an important functional role in coordinating the lymphatic contractile cycle [92] and in fine tuning lymphatic contractions to different levels of basal luminal flow [93]. Zhdanov and Zerbino reported ageing-related changes in morphology of various human lymphatic networks in the early 1960s [90, 94, 95]. They observed a reduction in the number of lymphatic capillaries (nonmuscular initial lymphatics) through all of the body and the presence of specific “varicose bulges,” which exist in muscular lymphatic vessels. It has also been reported that aged thoracic duct showed signs of lipid accumulation, thickening, and fibrosis [90, 96].
Recently, some authors reported changes in orientation and investiture of muscle cells in mesenteric lymphatic vessels in aged rats [90, 91]. It has been postulated that in elderly the decrease of accessory muscle elements surrounding lymphatic valve may limit the ability of lymphatic vessels to adapt their contractility to various preload/afterload challenges with subsequent formation of lymph stasis and potential spread of pathogens and immune cells in direction opposite to the direction of the normal lymph flow [90]. In addition, the thin-walled low muscle cells investiture zones in aged rats may be transformed to aneurysm-like formations “varicose bulges”, which can be ideal places for formation of low-velocity turbulent lymph flow and accumulation of various molecules, pathogens, and cancer cells [90]. Some studies reported a reduced lymph flow in aged animals in vivo [97, 98]. Ageing severely altered contractility of the toracic duct through weakening of lymphatic contractions and complete depletion of their shear/nitric oxide (NO)-dependent regulation [98]. It has been demonstrated that ageing severely altered NO-dependent regulation of thoracic duct contractions with an impaired eNOS function and an ageing-associated shear-independent NO release in the duct due to iNOS activation [98]. Non-specific nitric oxide synthase (NOS) blockade restored the contraction [98]. These findings provided functional consequences of ageing in lymphatic contractility and the dysfunctional responses of smooth muscle cells and endothelium in ageing-induced alterations [98].
Age-related changes of end-organ microvasculature
As a consequence of the age-related alterations in the expression and/or activity of eNOS, upregulation of iNOS, increased formation of ROS and ONOO-, and extracellular matrix remodelling, vasodilatory function is impaired and an excessive capillary pressure with consequent hyperfiltration, protein leakage, edema formation and tissue damage occur. In small arteries and arterioles, which have a relative higher wall thickness, changes in tone and circumferential shortening have an enhanced effect on lumen diameter, resulting in a blood flow decline in many organs [7]. We describe the main alterations that characterise the age-related end-organ damage.
Brain
Cognitive dysfunction from lower perfusion and microvascular fibrohyalinosis is the most common type of microvascular damage in the elderly [99]. Atherosclerosis in elderly people also coincides with massive microvascular fibrosis, which contributes to the development of white matter lesions, myelin rarefaction or demyelination, gliosis, apoptosis and regressive astrocytic changes [99–101]. Thickening of small vessels was associated with diffuse white matter lesions in elderly [102]. Reduced pericyte–endothelial cell contact also occurs [86].
Brain arteriolosclerosis is a subtype of cerebrovascular pathology characterised by concentrically thickened arterioles due to an altered proliferation of smooth muscle cells and excessive extracellular matrix deposition [103], as also shown in our histological study (Figure 3). Cerebral amyloid angiopathy (CAA) is another microvascular pathology associated with ageing and results from deposition of β-amyloid in the media and adventitia of small arteries and capillaries of the leptomeninges and cerebral cortex and is a major cause of lobar intracerebral hemorrhage and cognitive impairment in the elderly [104]. CAA is present in nearly all brains with Alzheimer disease, suggesting a common β-amyloid-based pathogenesis for these diseases. However, despite the close molecular relationship between the two diseases, CAA remains a clinically distinct entity from Alzheimer disease [104]. The accelerated β-amyloid vascular deposition in CAA seems to be caused by a transcriptional deregulation of the lipoprotein receptor LRP in VSMCs due to overexpression of the transcription factors: serum response factor (SRF) and myocardin [105]. In addition, SRF and myocardin may also regulate contractile proteins in VSMCs, thus altering normal vessel physiology [106].
Liver
Age-related changes in the human hepatic sinusoidal endothelium, termed pseudocapillarisation, have been recently described and they contribute to the impairment of hepatic function [107]. Blood clearance of a variety of waste macromolecules takes place in liver sinusoidal endothelial cells (SECs) [108]. These cells are unique endothelial cells in both their architecture and their function. The sinusoids are the exchange vessels of the liver, and the SECs are distinguished by extensive fenestrations organized into sieve plates, a lack of a basement membrane, and low junctional expression of CD31 [108]. The SEC architecture, including open fenestrations and weak junctional association between cells, provides a dynamic filtration system with low perfusion pressure that enables nutrients and macromolecular waste to pass freely to hepatocytes for efficient metabolism [108]. The maintenance of SEC phenotype is a critical process that requires both autocrine and paracrine cell signalling [108]. Recent studies indicate that fenestrations are maintained by constitutive VEGF-stimulated NO generation in SECs and surrounding cells [109]. In response to ageing [110], SECs dedifferentiate into a more regular endothelium, hence the term capillarisation or pseudocapillarisation. The hallmarks of capillarisation are SEC defenestration, development of a laminin-rich basement membrane, junctional expression of CD31 and protein nitration, in a mechanism involving NAD(P)H oxidase–generated ROS [108]. In addition, sinusoidal stellate cells are also induced to overexpress a laminin and collagen matrix that contributes to fibrosis [111].
In autoptic studies of older human subjects, independently from the presence of systemic diseases or hepatic pathologies, pseudocapillarisation occurs from increased peri-sinusoidal expression of von Willebrand’s factor, CD31 and collagen I and IV, resulting in a thickening and defenestration of the liver sinusoidal endothelium and deposition of basal lamina in the extracellular space of Disse [61, 107], as also shown in our histological study (Figure 4). In addition, it has been reported an endothelial upregulation of ICAM-1 [61]. Transmission electron microscopy study revealed a significant age-related thickening of the sinusoidal endothelium, with loss of fenestrations [61]. Loss of fenestrations leads to impaired transfer of lipoproteins from blood to hepatocytes. This provides a mechanism for impaired chylomicron remnant clearance and postprandial hyperlipidemia associated with old age [112].
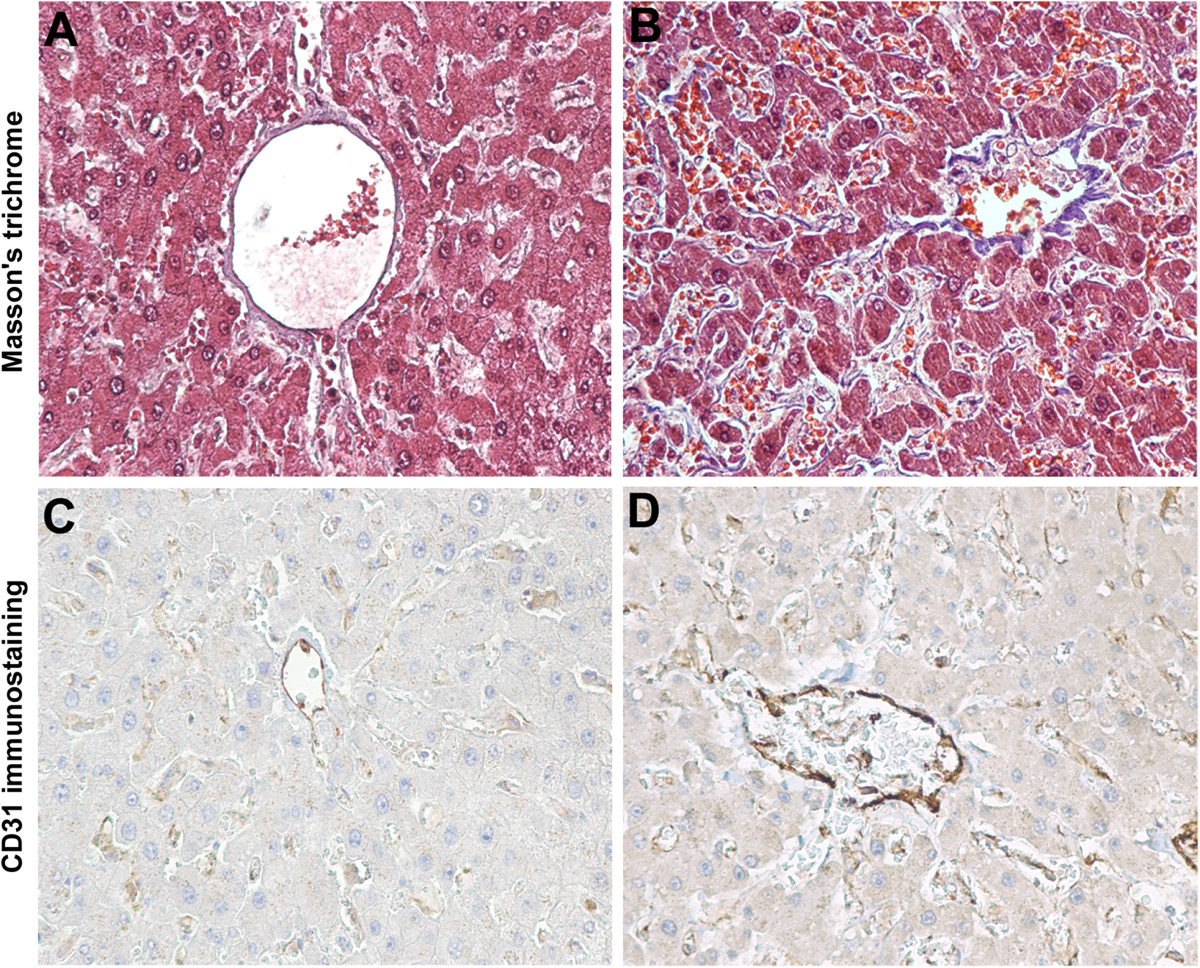
Figure 4
Figure 4 caption
Microscopic aspects of human liver pseudocapillarisation. Post-mortem (myocardial acute infarction) histology studies on paraffin-embedded sections (5 μm thick) of formalin-fixed liver tissue. Masson's trichrome staining shows the central vein and pericentral hepatocytes of young (A) and old liver (B) with perisinusoidal collagen deposition (blue staining). CD31 immunostaining of young (C) and old liver (D) with an increased sinusoidal protein expression. Magnification 20×.
Heart
Ageing is also associated with functional changes of the coronary microvasculature [113]. An important mechanism that contribute to the local regulation of myocardial blood flow is the flow (shear stress)–induced NO mediated dilatation of small coronary arteries and arterioles [114]; so ageing, that impairs NO synthesis/release in the endothelium (as described above), determines a vasodilatory dysfunction also in rat coronary arterioles [115]. It was also reported an increased breakdown of NO due to an augmented arteriolar production of O2- [116]. Moreover, in isolated coronary arterioles of old rats, with an impaired flow-induced dilatation, O2- and ONOO- production increased both in endothelial and VSMCs [36]. In addition, eNOS and SOD activity were impaired, whereas NAD(P)H oxidase and iNOS were upregulated. [36]. Aged human and rabbit small coronary vessels show a marked increase of myocardial interstitial collagen, with α-SMA and TGFβ-1 negative fibroblasts and VCAM-1 positive microvessels without macrophages [117, 118]; these findings support the close link between endothelial dysfunction and age-related fibrosis [117, 118]. The impaired coronary endothelial function may result in adverse clinical events because of the increased vascular and perivascular recruitment of neutrophils, macrophages, and platelets [119]. Taken together, these findings suggest that arteriolar changes, induced by ageing-related oxidative stress, impairs the vasoactive function of the coronary vessels in ageing.
Kidney and skin
With ageing, a degenerative process occurs with the appearance of glomerular lesions, as a thickening of the glomerular basement membrane and Bowman’s capsule [120], parallel to glomerulosclerosis, interstitial fibrosis and progressive proteinuria [121]. Biochemical studies evidenced the age-related increase of collagen and decrease in glycosaminoglycans, particularly of heparan sulphate [122]. Ultrastructural studies, conducted in our laboratory, documented a marked thickening of the glomerular basement membrane in old rats (Figure 5A-B). In addition, young rats perfused with cationized ferritin in vivo showed a regular distribution of these molecules, along the internal and external lamina rara of the glomerular basement membrane (Figure 5C). In the old rats, ferritin was present only along the internal lamina rara (Figure 5D), suggesting that the age-related loss of anionic charged of heparan sulphate molecules is responsible for age-related proteinuria, also reported in human. In the kidney of aged rats, the glomerular and peritubular capillary loss correlates with alterations in VEGF and TSP-1 expression and also with the development of glomerulosclerosis and tubulointerstitial fibrosis, suggesting an impaired angiogenesis associated with progressive loss in renal microvasculature [62]. The mechanism of capillary loss in aged kidney has not been fully understood. Angiostatin is a potent inhibitor of angiogenesis in vivo. In aged rats angiostatin production is increased, as well as the activity of cathepsin D, the enzyme for angiostatin production [123]. In addition, NO availability is decreased and cathepsin D activated, suggesting a possible correlation between the increase of angiostatin production, capillary loss and interstitial damage in aged rat kidney [123]. NOS inhibition by L-NAME produced a stronger vasoconstriction in renal vessels of old compared with young rats [124, 125], suggesting that endogenous NO production is necessary for the control of renal circulation. Moreover, post-mortem angiograms and histology studies, in elderly, showed wall thickening and narrowing of the vascular lumen of afferent arterioles, an alteration mainly depending on VSMC proliferation [126].
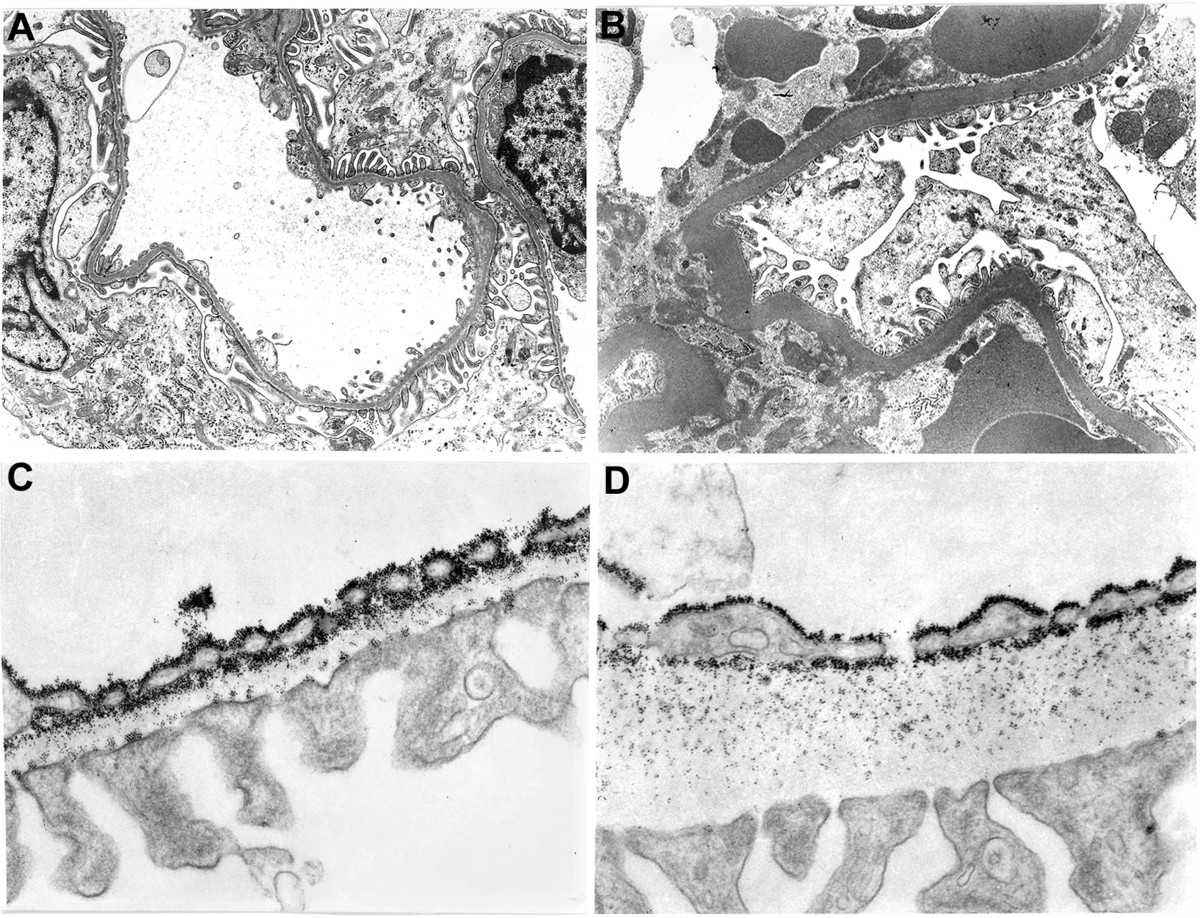
Figure 5
Figure 5 caption
Ultrastructural aspects of age-related changes in rat kidney microvessels. Glomerular basement membrane of kidney in young (A) and old rat (B), that shows the characteristic thickening of capillary wall. Magnification 5000×. Cationized ferritin distribution on glomerular basement membrane of young (C) and old rat kidney (D). Magnification 30000×.
Tubulointerstitial fibrosis, in aged rats, was characterised by tubular injury and focal tubular cell proliferation, myofibroblast activation, macrophage infiltration with increased immunostaining for the adhesive proteins osteopontin and ICAM-1, and collagen IV deposition, as well as a decrease in eNOS expression in peritubular capillaries [127]. In addition, it has been reported that ageing induced oxidative stress in kidney and the attenuation of redox status can ameliorate microvascular function [128]. Renal oxidative stress was associated with an increase in ONOO−, NO and ROS levels, as well as iNOS activity [129]. Treatment with an antioxidant reduced the age-related renal dysfunction [129]. Moreover, in aged rats, NF-κB activation has been reported to contribute to the accumulation of oxidative stress [130].
Structural and functional alterations of the skin during the ageing process are due to some complex mechanisms, determined by intrinsic and extrinsic factors, which act synergistically [131]. Collagen fibers become thinner and change their aspect; in the deep dermis they become more fibrous. Thickened microvessels can be recognised by the increased intensity of the vascular PAS positive-diastase resistant staining, and by the perivascular collagen deposition (Figure 6). Elastic fibers show the tendency of fragmentation, with a pathological assembly [131, 132]. With ageing, a progressive reduction of dermis vasculature is present, due to a reduction in the number and size of vascular vessels [131]. Age-related decrease in the number of dermal blood vessels is suggested to be due to an impairment of VEGF signalling [133]. In addition, it has been reported that eNOS activity is required for full expression of reflex cutaneous vasodilation, and its impairment in aged skin is associated with alterations in NO signalling [134], increase of oxidative stress and upregulation of arginase [135].
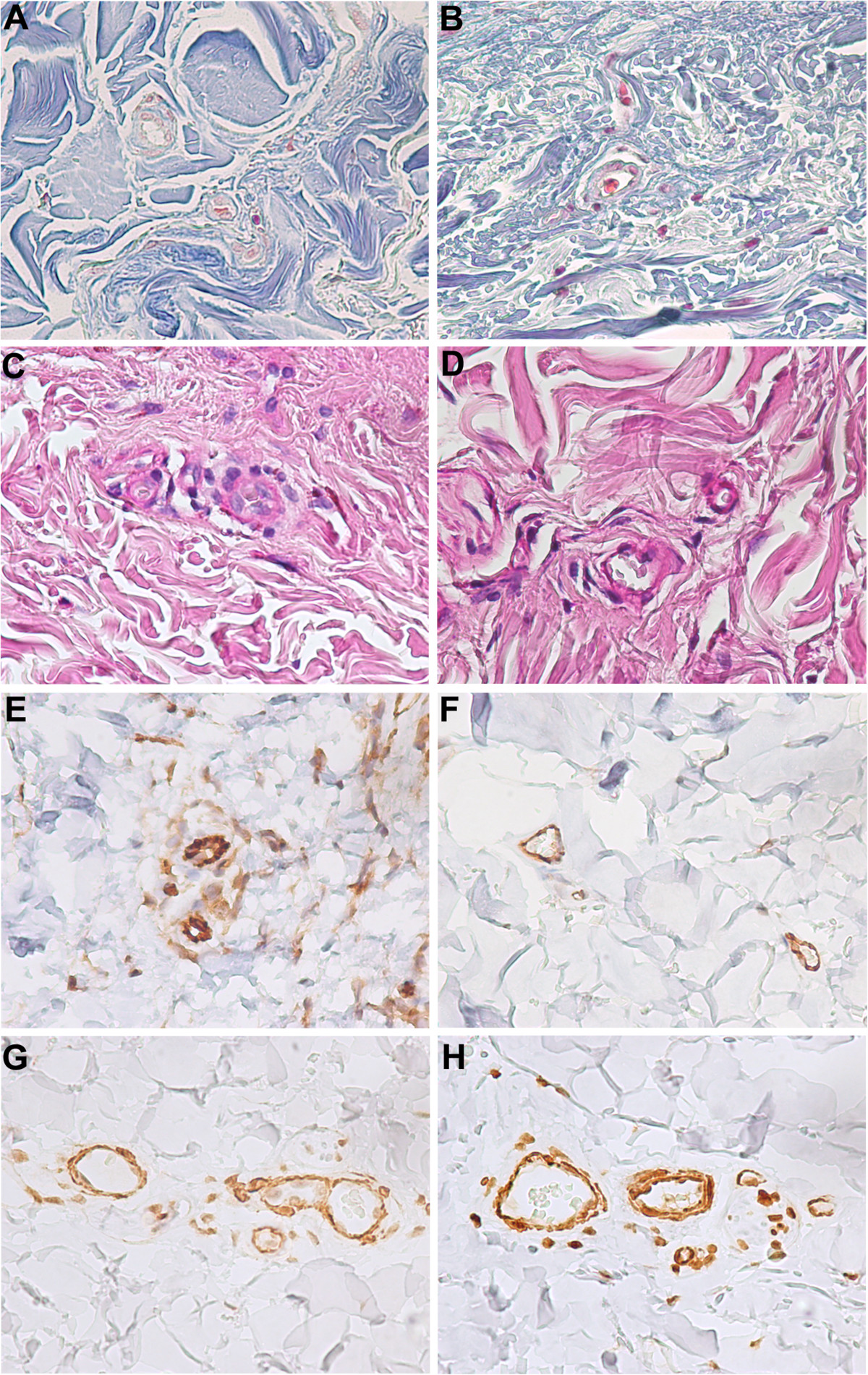
Figure 6
Figure 6 caption
Ageing in skin microcirculation. Histology studies on paraffin-embedded sections (5 μm thick) of formalin-fixed skin of healthy subjects. Masson's trichrome staining shows collagen distribution (blue staining), around microvessels, in young (A) and old dermal skin (B). PAS staining shows hyaline deposits (pink staining), around microvessels, in young (C) and old dermal skin (D). CD31 immunostaining of young (E) and old dermal skin (F) showing the descrease of capillaries associated with ageing process. α-SMA immunostaining of young (G) and old dermal skin (H) showing the proliferation of VSMCs around aged microvessels. Magnification 40×.
Therapeutic targeting of microvascular ageing
Being assumed that microvascular dysfunction plays a key role in age-related end-organ failure, several therapeutic attempts have been suggested. We summarised the most diffuse anti-atherosclerotic and anti-ischemic treatments and more anti-ageing innovative strategies.
Changes of lifestyle, anti-atherosclerotic and anti-ischemic treatments
Due to a high burden of cardiac risk factors and coronary atherosclerosis in subjects with angina and no obstructive coronary artery disease, lifestyle changes to modify risk factors are fundamental [136, 137]. Cardiac rehabilitation is recommended for those patients who have limited physical activity; increased exercise capacity is related to the amelioration of atherosclerotic disease symptoms [138]. Statins may improve endothelial function by lipid-independent anti-inflammatory and antioxidant properties and the capacity to restore microvascular NO availability [139]. Angiotensin-converting enzyme inhibitors as well as angiotensin-renin blockers [140] have been shown to improve endothelium-dependent relaxation of coronary arteries by increasing NO availability [141]. Upregulation of arginase has emerged as an important factor contributing to reduce NO production by competing with endothelial NO synthase for the common precursor substrate L-arginine [142]. Arginase inhibitors may induce long-term improvement of microvascular function and limitation of myocardial injury following ischaemia–reperfusion [143].
Antioxidant therapy
Some works focused the attention on antioxidant agents that can prevent or reduce the progression of end-organ microvascular dysfunction [144]. Antioxidants and free radical scavengers such as N-acetyl-cysteine (NAC), ascorbic acid and Propionyl-L-carnitine (PLC) showed a clinical efficacy in patients with endothelial dysfunction [145–149]. NAC, a derivative of cysteine, and ascorbic acid induced beneficial effects on oxidative stress and vascular dysfunction [145–147]. PLC is an ester of L-carnitine, that is required for the transport of fatty acids into the mitochondria [150]. PLC has been reported to modulate NF-kB activity in vascular cells [151] and to reduce age-related microvascular dysfunction and myocardial remodelling, including adhesion molecule expression [152]. In addition, it has been reported that PLC counteracts membrane lipid peroxidation and reduces post-ischemic endothelial dysfunction [153, 154].
Ascorbate is essential for normal endothelial function [155] and prevents microvascular dysfunction and H2O2-mediated injury in cultured microvascular endothelial cells [144]. Other natural substances, such as aged garlic extract and resveratrol, have been documented to minimise oxidative stress and to stimulate endothelial NO generation, suggesting that antioxidant regimens can be efficacy to counteract adverse clinical effects of age-related microvascular endothelial dysfunction [74, 75, 156]. In
Novel anti-inflammatory therapies
Vascular ageing is associated with deregulation of TNF-α expression [36, 159]. TNF-α is a master regulator of vascular inflammatory cytokines, chemokines and adhesion molecules. TNF-α plasma level increases with ageing and correlates with morbidity and mortality in the elderly patients [160, 161]. Consequently an anti-TNF-α treatment (i.e., with etanercept, which binds and inactivates TNF-α) may exert vasoprotective effects, including a reduction of endothelial cell apoptosis and the downregulation of NAD(P)H oxidases activity [162]. Pharmacological inhibition of the poly(ADP-ribose) polymerase (PARP) pathway also represents a novel therapeutic target to improve ageing-associated cardiovascular dysfunction [163].
Conclusions
Ageing elicits several structural and functional changes in the microvasculature. Reactive oxygen species and the concomitant oxidative and nitrosative stress play an important role in the process of ageing-related microvascular dysfunction, affecting vascular function as well as signalling transduction and gene expression. Although a significant progress has been achieved in describing the intrinsic age-related alterations of microvascular function, the age-related decline in endogenous antioxidant mechanisms, angiogenesis, endothelium-dependent vasodilation and microvascular permeability remains to be fully assessed. Increased knowledge may lead to new therapies targeting microvascular dysfunction and to improve clinical outcome. A key observation is that new therapeutic opportunities aimed to favour microvascular function are also associated with ameliorated organ function. An appropriate control of ageing process, in particular of oxidative stress, can clarify the efficacy of many pharmacological or nutritional approaches in order to delay the onset of age-dependent microvascular disease.
Acknowledgments
We thank Dr Sabrina Cappelli and Dr Antonio Volpe for their technical work.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Authors’ original file for figure 1
Authors’ original file for figure 2
Authors’ original file for figure 3
Authors’ original file for figure 4
Authors’ original file for figure 5
Authors’ original file for figure 6
References
- Age-related endothelial dysfunction with respect to nitric oxide, endothelium-derived hyperpolarizing factor and cyclooxygenase products. Physiol Res. 2000;49:11-18.
- Expert consensus document on arterial stiffness: methodological issues and clinical applications. Eur Heart J. 2006;27:2588-2605.
- Non-insulin-dependent diabetes mellitus and fasting glucose and insulin concentrations are associated with arterial stiffness indexes. The ARIC Study. Atherosclerosis Risk in Communities Study. Circulation. 1995;91:1432-1443.
- A definition of advanced types of atherosclerotic lesions and a histological classification of atherosclerosis. A report from the Committee on Vascular Lesions of the Council on Arteriosclerosis, American Heart Association. Arterioscler Thromb Vasc Biol. 1995;15:1512-1531.
- Age-related increase of stem marker expression influences vascular smooth muscle cell properties. Atherosclerosis. 2012;224:51-57.
- Hypertension and microvascular remodelling. Cardiovasc Res. 2008;78:274-285.
- Effects of central arterial aging on the structure and function of the peripheral vasculature: implications for end-organ damage. J Appl Physiol (1985). 2008;105:1652-1660.
- Human endothelial function and microvascular ageing. Exp Physiol. 2009;94:311-316.
- Microcirculation in hypertension: a new target for treatment?. Circulation. 2001;104:735-740.
- Microvascular dysfunction: a potential pathophysiological role in the metabolic syndrome. Hypertension. 2007;50:204-211.
- The physiology of the lymphatic system. Adv Drug Deliv Rev. 2001;50:3-20.
- Cross talk between mitochondria and NADPH oxidases. Free Radic Biol Med. 2011;51:1289-1301.
- Redox signaling, vascular function, and hypertension. Antioxid Redox Signal. 2008;10:1045-1059.
- Endothelial cell superoxide generation: regulation and relevance for cardiovascular pathophysiology. Am J Physiol Regul Integr Comp Physiol. 2004;287:R1014-R1030.
- Role of oxidative modifications in atherosclerosis. Physiol Rev. 2004;84:1381-1478.
- Redox signalling in vascular responses to shear and stretch. Cardiovasc Res. 2006;71:269-279.
- Vascular aging in the longest-living rodent, the naked mole rat. Am J Physiol Heart Circ Physiol. 2007;293:H919-H927.
- Endothelial aging. Cardiovasc Res. 2005;66:286-294.
- Relationship between antioxidants, prooxidants, and the aging process. Ann N Y Acad Sci. 1992;663:74-84.
- Age-dependent endothelial nitric oxide synthase uncoupling in pulmonary arteries of endoglin heterozygous mice. Am J Physiol Lung Cell Mol Physiol. 2009;297:L1170-L1178.
- 190 Nox2 activation and oxidative damage of cerebral vasculature and locomotor function in ageing mice. Heart. 2014;100(Suppl 3):A105-A106.
- Deletion of the activated protein-1 transcription factor JunD induces oxidative stress and accelerates age-related endothelial dysfunction. Circulation. 2013;127:1229-1240.
- Protection against vascular aging in Nox2-deficient mice: Impact on endothelial progenitor cells and reparative neovascularization. Atherosclerosis. 2012;223:122-129.
- Upregulation of superoxide dismutase and nitric oxide synthase mediates the apoptosis-suppressive effects of shear stress on endothelial cells. Arterioscler Thromb Vasc Biol. 1999;19:656-664.
- NAD(P)H oxidase-derived reactive oxygen species contribute to age-related impairments of endothelium-dependent dilation in rat soleus feed arteries. J Appl Physiol (1985). 2011;110:1171-1180.
- Oxidative stress and endothelial dysfunction in pulmonary arteries of aged rats. Am J Physiol Heart Circ Physiol. 2010;298:H346-H351.
- Age-related changes in renal expression of oxidant and antioxidant enzymes and oxidative stress markers in male SHR and WKY rats. Exp Gerontol. 2011;46:468-474.
- Reversal of persistent fibrosis in aging by targeting Nox4-Nrf2 redox imbalance. Sci Transl Med. 2014;6:231ra47-.
- Nitric oxide synthases: regulation and function. Eur Heart J. 2012;33:829-837.
- Enhanced peroxynitrite formation is associated with vascular aging. J Exp Med. 2000;192:1731-1744.
- Kinetics of superoxide dismutase- and iron-catalyzed nitration of phenolics by peroxynitrite. Arch Biochem Biophys. 1992;298:438-445.
- Quantitation of nitrotyrosine levels in lung sections of patients and animals with acute lung injury. J Clin Invest. 1994;94:2407-2413.
- Tyrosine nitration as a mechanism of selective inactivation of prostacyclin synthase by peroxynitrite. Biol Chem. 1997;378:707-713.
- Accumulation of nitrotyrosine on the SERCA2a isoform of SR Ca-ATPase of rat skeletal muscle during aging: a peroxynitrite-mediated process?. FEBS Lett. 1996;379:286-290.
- Vasodilator dysfunction in aged spontaneously hypertensive rats: changes in NO synthase III and soluble guanylyl cyclase expression, and in superoxide anion production. Cardiovasc Res. 1998;37:772-779.
- Aging-induced phenotypic changes and oxidative stress impair coronary arteriolar function. Circ Res. 2002;90:1159-1166.
- Endothelium-restricted overexpression of human endothelin-1 causes vascular remodeling and endothelial dysfunction. Circulation. 2004;110:2233-2240.
- Endothelin-1 decreases endothelial NOS expression and activity through ETA receptor-mediated generation of hydrogen peroxide. Am J Physiol Lung Cell Molecol Physiol. 2005;288:L480-L487.
- A role for mitochondrial oxidants in stress-induced premature senescence of human vascular smooth muscle cells. Redox Biol. 2013;1:411-417.
- Angiotensin II induces human astrocyte senescence through reactive oxygen species production. Hypertens Res. 2011;34:479-483.
- Immunohistochemical and molecular characterization of the differential response of the rat mesenteric microvasculature to angiotensin-II infusion. J Vascualr Res. 1996;33:195-208.
- Nitric oxide inhibition induces early activation of type I collagen gene in renal resistance vessels and glomeruli in transgenic mice. Role of endothelin. J Clin Invest. 1998;101:2780-2789.
- Impaired wound repair and delayed angiogenesis in aged mice. Lab Invest. 1999;79:1479-1487.
- Involvement of increased arginase activity in impaired cavernous relaxation with aging in the rabbit. J Urolol. 2004;172:369-373.
- Age-dependent impairment of angiogenesis. Circulation. 1999;99:111-120.
- Inhibited angiogenesis in aging: a role for TIMP-2. J Gerontol Ser A Biol Sci Med Sci. 2003;58:B798-B805.
- Intermittent high glucose enhances ICAM-1, VCAM-1 and E-selectin expression in human umbilical vein endothelial cells in culture: the distinct role of protein kinase C and mitochondrial superoxide production. Atherosclerosis. 2005;183:259-267.
- Inflammatory biomarkers in coronary artery disease. J Cardiol. 2009;53:317-333.
- Role of NFkappaB in age-related vascular endothelial dysfunction in humans. Aging (Albany NY). 2009;1:678-680.
- Aging influences development and progression of early aortic atherosclerotic lesions in cholesterol-fed rabbits. Arterioscler Thromb Vasc Biol. 2000;20:1123-1136.
- Microvascular disease and its role in the brain and cardiovascular system: a potential role for uric acid as a cardiorenal toxin. Nephrol Dial Transplant. 2011;26:430-437.
- Antioxidants and the bioactivity of endothelium-derived nitric oxide. Cardiovasc Res. 2000;47:426-435.
- Microvascular resuscitation as a therapeutic goal in severe sepsis. Crit Care (Lond Engl). 2005;9:S27-S32.
- Vascular smooth muscle and endothelial functions in aging. Ann New York Academi Sci. 2007;1100:353-360.
- Young adult bone marrow-derived endothelial precursor cells restore aging-impaired cardiac angiogenic function. Circ Res. 2002;90:E89-E93.
- Age-dependent depression in circulating endothelial progenitor cells in patients undergoing coronary artery bypass grafting. J Am Coll Cardiol. 2003;42:2073-2080.
- Persistent dysfunction of regenerated endothelium after balloon angioplasty of rabbit iliac artery. Circulation. 1990;81:1667-1679.
- Phenotypic and functional changes in regenerated porcine coronary endothelial cells: increased uptake of modified LDL and reduced production of NO. Circ Res. 2000;86:854-861.
- Ultrastructural and quantitative age-related changes in capillaries of the dorsal lateral geniculate nucleus. Brain Res Bull. 2004;64:145-153.
- Morphometric evaluation of capillaries in different laminae of rat cerebral cortex by automatic image analysis: changes during development and aging. Adv Neurol. 1978;20:1-9.
- Age-related pseudocapillarization of the human liver. J Pathol. 2003;200:112-117.
- Impaired angiogenesis in the aging kidney: vascular endothelial growth factor and thrombospondin-1 in renal disease. Am J Kidney Dis. 2001;37:601-611.
- Mechanisms of vascular aging: new perspectives. J Gerontol A Biol Sci Med Sci. 2010;65:1028-1041.
- Age-related changes in redox signaling and VSMC function. Antioxid Redox Signal. 2010;12:641-655.
- Aging, oxidative responses, and proliferative capacity in cultured mouse aortic smooth muscle cells. Am J Physiol Heart Circ Physiol. 2001;280:H2779-H2788.
- Modulation of oxidant and antioxidant enzyme expression and function in vascular cells. Hypertension. 2004;44:381-386.
- Aging influences multiple incidices of oxidative stress in the aortic media of the Fischer 344/NNiaxBrown Norway/BiNia rat. Free Radic Res. 2006;40:185-197.
- Upregulation of Nox1 in vascular smooth muscle leads to impaired endothelium-dependent relaxation via eNOS uncoupling. Am J Physiol Heart Circ Physiol. 2010;299:H673-H679.
- Calcium-dependent NOX5 nicotinamide adenine dinucleotide phosphate oxidase contributes to vascular oxidative stress in human coronary artery disease. J Am Coll Cardiol. 2008;52:1803-1809.
- Nox2 as a potential target of mitochondrial superoxide and its role in endothelial oxidative stress. Am J Physiol Heart Circ Physiol. 2013;305:H1131-H1140.
- Role of NADH/NADPH oxidase-derived H2O2 in angiotensin II-induced vascular hypertrophy. Hypertension. 1998;32:488-495.
- Cross-talk between aldosterone and angiotensin II in vascular smooth muscle cell senescence. Cardiovasc Res. 2007;76:506-516.
- Increased expression of matrix metalloproteinase-2 in the thickened intima of aged rats. Hypertension. 1999;33:116-123.
- Increased mitochondrial H2O2 production promotes endothelial NF-kappaB activation in aged rat arteries. Am J Physiol Heart Circ Physiol. 2007;293:H37-H47.
- Resveratrol delays age-related deterioration and mimics transcriptional aspects of dietary restriction without extending life span. Cell Metab. 2008;8:157-168.
- Vascular cell senescence: contribution to atherosclerosis. Circ Res. 2007;100:15-26.
- β-adrenergic receptor responsiveness in aging heart and clinical implications. Front Physiol. 2013;4:396-.
- Aging, smooth muscle cells and vascular pathobiology: implications for atherosclerosis. Atherosclerosis. 2006;188:221-230.
- Age-related changes in the signaling and function of vascular smooth muscle cells. Exp Gerontol. 1999;34:549-557.
- Role of the extracellular matrix in age-related modifications of the rat aorta. Ultrastructural, morphometric, and enzymatic evaluations. Arterioscler Thromb. 1992;12:1008-1016.
- Cardiovascular regulatory mechanisms in advanced age. Physiol Rev. 1993;73:413-467.
- The vascular smooth muscle cell in arterial pathology: a cell that can take on multiple roles. Cardiovasc Res. 2012;95:194-204.
- Inhibition of capillary endothelial cell growth by pericytes and smooth muscle cells. J Cell Biol. 1987;105:1455-1462.
- Pericyte loss and microaneurysm formation in PDGF-B-deficient mice. Science. 1997;277:242-245.
- Pericytes in the microvasculature. Cardiovasc Res. 1996;32:687-698.
- Altered pericyte-endothelial relations in the rat retina during aging: implications for vessel stability. Neurobiol Aging. 2006;27:1838-1847.
- Cellular senescence, vascular disease, and aging: part 2 of a 2-part review: clinical vascular disease in the elderly. Circulation. 2011;123:1900-1910.
- A quantitative analysis of blood–brain barrier ultrastructure in the aging human. Microvasc Res. 1987;33:270-282.
- Type-1 pericytes participate in fibrous tissue deposition in aged skeletal muscle. Am J Physiol Cell Physiol. 2013;305:C1098-C1113.
- Lymphatic muscle cells in rat mesenteric lymphatic vessels of various ages. Lymphat Res Biol. 2013;11:35-42.
- Basic mechanisms controlling lymph transport in the mesenteric lymphatic net. Ann New York Acad Sci. 2010;1207:E16-E20.
- Phasic contractions of rat mesenteric lymphatics increase basal and phasic nitric oxide generation in vivo. Am J Physiol Heart Circ Physiol. 2009;297:H1319-H1328.
- Regional variations of contractile activity in isolated rat lymphatics. Microcirculation. 2004;11:477-492.
- Senile changes in the lymphatic capillaries and vessels. Arkh Anat Gistol Embriol. 1960;39:24-36.
- Senile changes in the outflow lymphatic vessels. Arkh Anat Gistol Embriol. 1960;39:37-42.
- Changes in the structure of the wall of the human thoracic duct in relation to atherosclerosis and age. Morphol Igazsagugyi Orv Sz. 1976;16:43-47.
- Lymphatic absorption of retinol in young, mature, and old rats: influence of dietary restriction. FASEB J. 1996;10:1085-1090.
- Age-related alterations of active pumping mechanisms in rat thoracic duct. Microcirculation. 2007;14:827-839.
- Pathologies and pathological mechanisms for white matter hyperintensities in depression. Ann N Y Acad Sci. 2002;977:333-339.
- Apoptosis in leukoaraiosis. AJNR Am J Neuroradiol. 2000;21:79-82.
- Apoptosis of astrocytes with enhanced lysosomal activity and oligodendrocytes in white matter lesions in Alzheimer's disease. Neuropathol Appl Neurobiol. 2002;28:238-251.
- Age-related microvascular degeneration in the human cerebral periventricular white matter. Acta Neuropathol. 2006;111:150-157.
- Arteriolosclerosis that affects multiple brain regions is linked to hippocampal sclerosis of ageing. Brain J Neurol. 2014;137:255-267.
- Cerebral amyloid angiopathy in the elderly. Ann Neurol. 2011;70:871-880.
- SRF and myocardin regulate LRP-mediated amyloid-beta clearance in brain vascular cells. Nat Cell Biol. 2009;11:143-153.
- Serum response factor and myocardin mediate arterial hypercontractility and cerebral blood flow dysregulation in Alzheimer's phenotype. Proc Natl Acad Sci U S A. 2007;104:823-828.
- Capillarization of hepatic sinusoid by liver endothelial cell-reactive autoantibodies in patients with cirrhosis and chronic hepatitis. Am J Pathol. 2003;163:1275-1289.
- Arsenic-stimulated liver sinusoidal capillarization in mice requires NADPH oxidase-generated superoxide. J Clin Invest. 2008;118:3980-3989.
- Rat liver sinusoidal endothelial cell phenotype is maintained by paracrine and autocrine regulation. Am J Physiol Gastrointest Liver Physiol. 2004;287:G757-G763.
- Age-related changes in the hepatic sinusoidal endothelium impede lipoprotein transfer in the rat. Hepatology. 2005;42:1349-1354.
- Sinusoidal endothelial cells prevent rat stellate cell activation and promote reversion to quiescence. Hepatology. 2008;48:920-930.
- Old age and the hepatic sinusoid. Anat Rec (Hoboken). 2008;291:672-683.
- Role of capillaries in determining CBF reserve: new insights using myocardial contrast echocardiography. Am J Physiol. 1999;277:H2363-H2372.
- Endothelium-dependent, flow-induced dilation of isolated coronary arterioles. Am J Physiol. 1990;259:H1063-H1070.
- Expression of constitutive and inducible nitric oxide synthases in the vascular wall of young and aging rats. Circ Res. 1998;83:279-286.
- Superoxide excess in hypertension and aging: a common cause of endothelial dysfunction. Hypertension. 2001;37:529-534.
- Role of ageing and coronary atherosclerosis in the development of cardiac fibrosis in the rabbit. Cardiovasc Res. 2004;64:544-552.
- Alpha-actin isoform distribution in normal and failing human heart: a morphological, morphometric, and biochemical study. J Pathol. 2003;199:387-397.
- Aging-associated endothelial dysfunction in humans is reversed by L-arginine. J Am Coll Cardiol. 1996;28:1796-1804.
- Spontaneous glomerular sclerosis in aging Sprague–Dawley rats. II. Ultrastructural studies. Am J Pathol. 1980;98:339-356.
- Spontaneous glomerular sclerosis in aging Sprague–Dawley rats. I. Lesions associated with mesangial IgM deposits. Am J Pathol. 1976;85:277-302.
- Changes in the glomerular filtration barrier during aging in rats. C R Acad Sci III. 1984;299:379-382.
- Angiostatin production increases in response to decreased nitric oxide in aging rat kidney. Lab Invest. 2013;93:334-343.
- Basal and stimulated nitric oxide in control of kidney function in the aging rat. Am J Physiol. 1999;272:R1747-R1753.
- Role of nitric oxide-related mechanisms in renal function in ageing rats. Nephrol Dialisis Transplant. 1998;13:594-601.
- Blood vessels and the aging kidney. Nephron Exp Nephrol. 2005;101:e95-e99.
- Tubulointerstitial disease in aging: evidence for underlying peritubular capillary damage, a potential role for renal ischemia. J Am Soc Nephrol. 1998;9:231-242.
- The essential role of FoxO6 phosphorylation in aging and calorie restriction. Age (Dordr). 2014;36:9679-.
- Down-regulation of oxidative stress and COX-2 and iNOS expressions by dimethyl lithospermate in aged rat kidney. Arch Pharm Res. 2014;37:1032-1038.
- Oxidative stress induces inactivation of protein phosphatase 2A, promoting proinflammatory NF-κB in aged rat kidney. Free Radic Biol Med. 2013;61C:206-217.
- The process of ageing reflected by histological changes in the skin. Rom J Morphol Embryol. 2013;54:797-804.
- Studies in cutaneous aging: II. The microvasculature. J Invest Dermatol. 1982;78:444-448.
- Age-related changes in angiogenesis in human dermis. Exp Gerontol. 2014;55:143-151.
- Decreased active vasodilator sensitivity in aged skin. Am J Physiol. 1997;272:H1609-H1614.
- Local tetrahydrobiopterin administration augments reflex cutaneous vasodilation through nitric oxide-dependent mechanisms in aged human skin. J Appl Physiol. 2012;112:791-797.
- Aging and vascular endothelial function in humans. Clin Sci (Lond). 2011;120:357-375.
- Angina with "normal" coronary arteries: a changing philosophy. JAMA. 2005;293:477-484.
- Physical training in Syndrome X: physical training counteracts deconditioning and pain in Syndrome X. J Am Coll Cardiol. 2000;36:1619-1625.
- Endothelial dysfunction: a marker of atherosclerotic risk. Arterioscler Thromb Vasc Biol. 2003;23:168-175.
- Telmisartan, an angiotensin II type 1 receptor blocker, improves coronary microcirculation and insulin resistance among essential hypertensive patients without left ventricular hypertrophy. Hypertens Res. 2008;31:615-622.
- ACE inhibitors and statins acutely improve endothelial dysfunction of human coronary arterioles. Am J Physiol Heart Circ Physiol. 2004;286:H1425-H1432.
- Arginase: a critical regulator of nitric oxide synthesis and vascular function. Clin Exp Pharmacol Physiol. 2007;34:906-911.
- Arginase inhibition improves coronary microvascular function and reduces infarct size following ischaemia-reperfusion in a rat model. Acta Physiol (Oxf). 2013;208:172-179.
- Ascorbate prevents microvascular dysfunction in the skeletal muscle of the septic rat. J Appl Physiol (1985). 2001;90:795-803.
- Influence of N-acetylcysteine on ICAM-1 expression and IL-8 release from endothelial and epithelial cells. J Physiol Pharmacol. 2006;57:325-334.
- Antioxidant vitamins in atherosclerosis–animal experiments and clinical studies. Adv Clin Exp Med. 2012;21:115-123.
- Role of vitamin C in the function of the vascular endothelium. Antioxid Redox Signal. 2013;19:2068-2083.
- A systematic review to evaluate the effectiveness of carnitine supplementation in improving walking performance among individuals with intermittent claudication. Atherosclerosis. 2013;229:1-9.
- A systematic review and meta-analysis of propionyl-L-carnitine effects on exercise performance in patients with claudication. Vasc Med (Lond Engl). 2013;18:3-12.
- Carnitine–metabolism and functions. Physiol Rev. 1983;63:1420-1480.
- Propionyl-L-carnitine reduces proliferation and potentiates Bax-related apoptosis of aortic intimal smooth muscle cells by modulating nuclear factor-kappaB activity. J Biol Chem. 2007;282:4932-4942.
- Propionyl-L-carnitine prevents age-related myocardial remodeling in the rabbit. J Cardiovasc Pharmacol. 2007;50:168-175.
- Myocyte performance during evolution of myocardial infarction in rats: effects of propionyl-L-carnitine. Am J Physiol. 1995;268:H1702-H1713.
- Propionyl-L-carnitine improves postischemic blood flow recovery and arteriogenetic revascularization and reduces endothelial NADPH-oxidase 4-mediated superoxide production. Arterioscler Thromb Vasc Biol. 2010;30:426-435.
- Aortic wall damage in mice unable to synthesize ascorbic acid. Proc Natl Acad Sci U S A. 2000;97:841-846.
- Aged garlic extract improves homocysteine-induced endothelial dysfunction in macro- and microcirculation. J Nutr. 2006;136:750S-754S.
- Vasoprotective effects of resveratrol and SIRT1: attenuation of cigarette smoke-induced oxidative stress and proinflammatory phenotypic alterations. Am J Physiol Heart Circ Physiol. 2008;294:H2721-H2735.
- Resveratrol induces mitochondrial biogenesis in endothelial cells. Am J Physiol Heart Circ Physiol. 2009;297:H13-H20.
- Proinflammatory phenotype of coronary arteries promotes endothelial apoptosis in aging. Physiol Genomics. 2004;17:21-30.
- Ageing, tumour necrosis factor-alpha (TNF-alpha) and atherosclerosis. Clin Exp Immunol. 2000;121:255-260.
- Associations of elevated interleukin-6 and C-reactive protein levels with mortality in the elderly. Am J Med. 1999;106:506-512.
- Vasculoprotective effects of anti-tumor necrosis factor-alpha treatment in aging. Am J Pathol. 2007;170:388-398.
- A new, potent poly(ADP-ribose) polymerase inhibitor improves cardiac and vascular dysfunction associated with advanced aging. J Pharmacol Exp Ther. 2004;311:485-491.

The present article has been published in Vascular Cell journal by Publiverse Online S.R.L.